PALEONTOLÓGIA
JORDÁN FERENC
A kihalások története
A Földet benépesítô élôlények
sokféleségét a fajok képzôdése
és kihalása szabja meg. A kihalás olykor tömeges
méreteket ölt, mint napjainkban is. A jelenkori események
megértésének egyik kulcsa a múlt ismerete lehet.
Kérdés, hogyan lehetett és lehet biztonságban
túlélni. Az élôvilág története
során sok élôlény végleg eltûnt
már bolygónkról. A kihalások mindig hozzátartoztak
az evolúció krónikájához, de annyi faj
valószínûleg sosem pusztult ki, mint napjainkban. Írásom
célja, hogy egy naprakész, és a lehetôségekhez
mérten átfogó ismertetés mellett újszerû
nézôpontból mutassam be a kihalások történetét.
A fajok túlélési esélyét egyedeik
különféle életmûködéseinek megbízhatóságával
lehet jellemezni. Ha a gepárd nagy valószínûséggel
utoléri és leteríti az antilopot, zsákmányszerzô
viselkedése megbízható. Ez hozzájárul
ahhoz, hogy kisebb eséllyel pusztuljon ki, mert a kisebb-nagyobb
környezeti változásokat megbízhatóbb mûködés
mellett könnyebb tolerálni. A megbízhatóságelméleti
keret módot ad az amúgy sokszínû téma
egységes tárgyalására.
Kihalás (extinkció) alatt egy leszármazási
vonal megszakadását értjük. Elôfordulhat,
hogy egy faj geológiai idômértékkel mérve
gyorsan megváltozik és másik fajjá alakul.
Ilyenkor az ôsibb faj nyomai eltûnnek a késôbbi
fosszilis anyagból, de a leszármazási vonal nem szakad
meg. Ezt a jelenséget álkihalásnak (pszeudoextinkció)
nevezzük.
Nehéz elképzelni, milyen lenne ma az élôvilág,
ha nem történnének idônként kisebb-nagyobb
kihalások. A sokféleség valószínûleg
gyors, kezdetben exponenciális mértékben növekedett
volna. Késôbb talán telítési görbének
megfelelôen beállt volna egy állandó fajszám,
a fajképzôdés megszûnésével. Az
is lehet, hogy kis egyedszámú, nagyon erôsen specializálódott
fajok alakultak volna ki. Ha nem történtek volna kihalások,
az állatkertekben megcsodálhatnánk a kambriumi fura
lényeket, vagy mondjuk a dodót és a dinoszauruszokat
is.
A kihaláskutatás története
Sokáig azt gondolták a talált kövületekrôl,
hogy létezô, de még fel nem fedezett fajok elpusztult
tagjainak maradványai. A 18. század végére
azonban a természetbúvárok már bejárták
a Föld nagy részét és kezdtek gyanakodni, vajon
hogyan kerülhette el a figyelmüket a kardfogú tigris vagy
a mamut.
Franciaországban George Cuvier és tanítványai
azt hirdették, hogy az ôsmaradványok hatalmas katasztrófák
során kipusztult fajok emlékei. Ez volt a katasztrofista
iskola. Angliában Charles Lyell és késôbb
Darwin köre a
kövületekben szintén már kipusztult fajokat vélt
felismerni, de ôk a fajok kihalását fokozatos és
szórványos eseménynek tartották. Ez a korai
kihaláskutatás másik, gradualista iskolája,
mely a kihalások folytonosságát hirdette.
A modern kihaláskutatás századunk ötvenes-hatvanas
éveiben lendült fel, majd 1980-ban Alvarez és
társainak szenzációszámba menô cikke
adott nagy lökést a tudományterületnek. Ebben a
cikkben vetôdött fel elôször a dinoszauruszok kipusztulásának
magyarázataként a Földre becsapódó óriási
meteor lehetôsége.
Ma a fajok kihalását fôleg paleontológusok
és biológusok kutatják. A paleontológusok az
ôsmaradványok elemzését tartják a legbiztosabb
módszernek. A biológusok ezzel szemben egyrészt a
mai élôvilág megfigyelésébôl vonnak
le a múltra vonatkozó következtetéseket, másrészt
pedig matematikusokkal és fizikusokkal együttmûködve
igyekeznek modellezni a múltbeli eseményeket. Az igazán
komoly eredményeknek természetesen meg kell felelniük
mindegyik megközelítés elvárásainak.
A kihalások változatai
A földtörténet egyes korszakaiból különbözô
számú emlékünk maradt. A kriptozoikum körülbelül
570 millió éve ért véget. Neve arra utal, hogy
keveset tudunk róla, az akkoriban élt élôlények
ugyanis szilárd váz hiányában nehezen fosszilizálódtak.
Van persze néhány kivétel, például egysejtû
organizmusok nyomai Dél-Afrikából. Az elmúlt
570 millió év a fanerozoikum. A fosszilis anyag döntô
többsége ebbôl az idôszakból származik.
A kövületek minôsége és mennyisége
változó. Függ tértôl, idôtôl
és magától az élôlénytôl.
Sok lelet ôrzi például jura idôszaki tengeri
csigák emlékét. A fosszilis anyag kétségtelenül
a legközvetlenebb lehetôség a múlt vizsgálatára,
de mindig figyelembe kell venni a szelektivitását és
a szórványosságát. A kövületek többsége
nem feltétlenül a leggyakoribb, hanem a legkönnyebben
megmaradó fajokról árulkodik. Egy-egy élôlénycsoport
kielégítô elemzését és leírását
nehezíti a fennmaradt leletek csekély mennyisége,
mely például statisztikailag csak ritkán értékelhetô
ki.
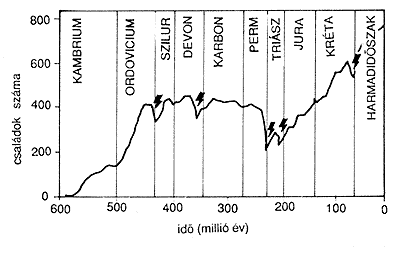
1. ábra. Az elmúlt 600 millió év során
így alakult a családok
száma. A fekete nyilak az öt legnagyobb kihalási eseményt
jelzik.
Ezek az ordovicium, a devon, a perm, a triász és a kréta
idôszak
végén történtek. Az adatok tengeri élôlényekre
vonatkoznak
(E. O. Wilson, 1992 nyomán). |
Az élet története során mindig pusztultak
ki fajok, de nem ugyanolyan mértékben. Egyes idôszakokban
tömeges kihalásokról lehet beszélni, eltérôen
a folyamatosan és nagyjából azonos mértékben
végbemenô háttérkihalási folyamatoktól.
A legnagyobb tömeges kihalások ideje az ordovicium, a devon,
a perm, a triász és a kréta idôszak végére
tehetô. Többek között ezekhez a jelentôs eseményekhez
igazították a földtörténeti korbeosztást;
az egyes korok végével való egybeesés tehát
nem a véletlen mûve. Ezek közül a legrégebbi,
az ordovicium végi tömeges kihalás kb. 440 millió
éve történt, a legutóbbi, a kréta végi
pedig kb. 65 millió éve. Egyesek más tömeges
kihalásokat is számon tartanak, például E.
O. WiIson a hatodik ilyen eseménynek tartja a mai kor minden
eddigit felülmúló kipusztulási hullámát.
Fontos, még el nem döntött kérdés, vajon
a tömeges kihalások minôségileg mások-e,
vagy csupán a háttérkihalás felerôsödésébôl
származnak. David Jablonski szerint a tömeges kihalások
alkalmával más törvényszerûségek
érvényesülnek. Ilyenkor nem számít a fajok
alkalmazkodottsága, megnô a véletlen szerepe. Ez remek
lehetôséget ad addig jelentéktelenebb csoportok gyors
kivirágzására. Ennek köszönhetô az
emlôsök mai uralma.
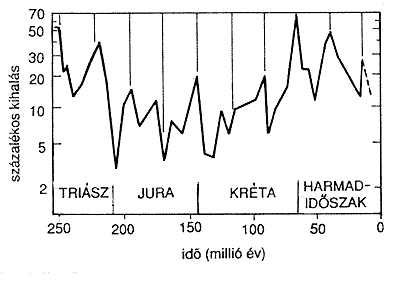
2. ábra. Az utolsó 250 millió év elemzése
alapján Raup
és Sepkoski a kihalások idôrendjében 26 millió
éves
periodicitást vél fölfedezni. Az ábra felsô
részén látható
függôleges csíkok távolsága ennek felel
meg
(D. M. Raup és J. Sepkoski, 1984 nyomán). |
Néhány kutató rendszerességet vél
felfedezni a nagyobb kihalási események idôrendjében.
David Raup és John Sepkoski, két neves chicagói
paleontológus szerint a számottevôbb kihalási
események ciklusideje 26 millió év. Eredményeiket
sokan bírálják; állítólag az
általuk figyelembe vett kihalások 75 százaléka
álkihalás. M. J. Benton szerint a nagyobb kihalási
idôszakok közt elte(t idô 20 és 60 millió
év között változik, így semmi értelme
periodicitásról beszélni.
A bioszféra diverzitásának változásáról
alkotott kép is változatos. Egyesek a diverzitás állandó
szintjérôl beszélnek, mások exponenciális
növekedésrôl. E. O. Wilson ahhoz a legnépesebb
csoporthoz tartozik, mely szerint a diverzitás lassan nô.
Ennek okát is keresik. Valószínûleg nagyjából
állandó a fajképzôdés mértéke,
míg az átlagos kihalási valószínûség
lassan csökken. Wilson megfogalmazása szerint a diverzitásváltozás
története a fajképzôdés (speciáció)
és a kihalás (extinkció) egyensúlyának
nyelvén íródik. A csökkenô átlagos
kihalási valószínûség egy elôremutatónak
mondható evolúciós trend eredménye is lehet,
de származhat pusztán abból is, hogy a fosszilis adat
egyre teljesebb (ha egy régebbi korszakból a hiányosabb
fosszilis anyag miatt egy rendszertani egység képviselôit
nem találjuk és emiatt a taxont kihaltnak véljük,
a kihalási rátát felbecsüljük).
Az egyes fajok kihalási valószínûsége
független a faj evolúciós múltjának hosszától.
Függ viszont az alkalmazkodottság mértékétôl,
ha nem túl erôs a történeti esetlegesség
(kontingencia). Ideális esetben az evolúciós siker
szigorúan jelzi az alkalmazkodottság mértékét,
de a valóságban mindig szerephez jut a véletlen is.
Az alkalmazkodottság egyik összetevôje a megbízható
mûködés. A leszármazási vonal túlélésének
megbízhatósága az élettevékenységek
sikeres elvégzésének valószínûsége.
A megbízhatóság mértéke különféle
módokon növelhetô, megbízhatósági
determinánsok beépítésével. Ilyen például
a redundancia (ivarsejtek száma), a raktározás
(zsírszövet), a javítás (regenerálódó
szövetek) vagy a tevékenységek optimális szervezése
(a rovartársadalmak mûködésében). Ha
az életmûködések alacsony megbízhatóságúak,
a faj kihalhat, például mert fokozottan érzékeny
a környezet apróbb változásaira is. A túl
nagy mértékû megbízhatóság is
hátrányos lehet. Egyrészt, mert a megbízhatósági
determinánsok költségesek. Aki túl sokat fektet
be a megbízhatóságba, más területen lemaradhat
és például a versengésben való lemaradás
révén kipusztulhat. Másrészt, mert a túlzott
mértékû megbízhatóság rugalmatlanságot
eredményez és változó környezetben ez
is kihaláshoz vezethet. Az élôlények tehát
életmûködéseik optimális szinten tartott
megbízhatóságával védekezhetnek a kihalás
ellen.
A kihalások szelektivitása más szempontok szerint
is jellemzô. Függ például az élôhelytôl.
A trópusokon általában nagyobb a kihalás mértéke.
Ez gyorsabb fajképzôdéssel párosul, így
a tömeges kihalások után a trópusi területek
forrásként szolgálhatnak más vidékek
benépesüléséhez is. Általánosan
is igaznak tûnik, hogy a nagyobb evolúciós intenzitású
idôszakokban és területeken a fajképzôdés
és a kihalás szintje is magasabb. Jablonski a kihalások
szelektivitásával kapcsolatban felsorol olyan jellegeket,
melyek elôsegíthetik egy faj fennmaradását.
Ilyen a jó elterjedési képesség, a nagy elterjedési
terület vagy a magas termelékenység. Friss eredmények
szerint a ma élô 9672 madárfaj közül 1111
veszélyeztetett. A veszélyeztetettség mértéke
erôsen korrelál a kis termékenységgel és
a nagy testmérettel. A nagyobb testû élôlények
kétszeresen veszélyeztetettebbek. A nagy testméret
ugyanis egyrészt hosszú generációs idôt
jelent, ami rugalmatlanságot okoz, ha a környezet elég
gyorsan változik. Másrészt pedig csak kis populációméretet
enged meg, mely kiteszi a populációt a véletlenszerû
ingadozások felerôsödésével járó
veszélyeknek. Ugyanakkor egy kis testû egyedekbôl álló
populációban sok szempontból elônyben vannak
a nagyobb testû egyedek. A testmérettel kapcsolatban tehát
megjegyezhetjük, hogy az egyed és a faj érdekei konfliktusban
vannak egymással.
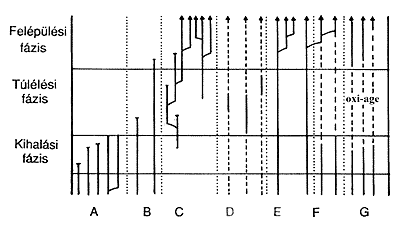
3. ábra. A tömeges kihalási idôszakok három
fázisa a kihalási,
a túlélési és a felépülési
fázis. A különbözô rendszertani
csoportok hétféleképpen viselkedhetnek egy ilyen kritikus
idôszakban. Bôvebb magyarázat a szövegben
(A. Hallam és P B. Wignall, 1997 nyomán). |
A tömeges kihalási idôszakok során az egyes
taxonok sorsa sokféle lehet. Antony Hallam angol geológus
a következô hét típusba sorolja ez alapján
a taxonokat. Az "A" csoport tagjai azok, amelyek a tömeges
kihalás során maguk is kihalnak. Az "E"-be tartoznak
azok, amelyek folyamatosan jelen vannak és végül túlélik
a kritikus idôszakot. A "B" csoport tagjai nagyrészt
kihalnak a kihalási hullám csúcsán, de a taxon
néhány faja csak valamivel késôbb pusztul ki.
A pörgekarúak (a tapogatókoszorúsok törzsének
egyik osztálya) nagyobb része például kihalt
a perm végén (250 millió éve), de sok csoportjuk
csak a triászban tûnt el végleg (200 millió
éve). A "C" csoport tagjai azok, amelyek a nagy kihalást
átvészelik, majd virágzásnak indulnak és
betöltik a megüresedett élettereket. Vannak, amelyek általában
kis egyedszámban fordulnak elô, de a kihalási idôszakokban
virágkorukat élik ("D" csoport). Az Earlandia nevû
foraminiferák például a devon és a perm végén
éltek tömegesen. A "G" csoport tagjai a kritikus
idôszak elôtt és után nagy számban képviseltetik
magukat, de a kritikus idôszakban szinte eltûnnek a Föld
színérôl. Végül pedig az "F"
csoportba tartoznak az Elvis-típusú taxonok. Ezek a tömeges
kihalás áldozataiként kihalnak, de késôbb
látszólag újra megjelennek. Valójában
más, túlélô élôlénycsoportok
imitálják ôket konvergens evolúció révén.
A konvergens evolúció eltérô fejlôdési
eredetû, de felépítésükben és mûködésükben
hasonló, homopláziás szervek kialakulását
jelenti. (Az elnevezés a Király imitátorainak emlékét
ôrzi.)
A kihalásokat magyarázó elméletek
A kihalások okára magyarázatot kínáló
elméleteknek négy csoportját mutatom be. Elöljáróban
érdemes végiggondolni, hogy ha elfogadjuk a kihalási
események szigorú periodicitását, kizárólagos
okot érdemes keresnünk, vagy legalábbis olyat, amely
erôsen domináns a többi felett. A bioszféra rendkívüli
bonyolultságát ismerve azonban sokkal valószínûbb,
hogy sok hatás együtteseként kell elképzelnünk
a globális változások okait.
A biológiai tényezôket fôleg a folyamatos
háttérkihalások okaként ismerik el. Az azonos
életközösségben élô élôlények
közös evolúciós múltjuk miatt egymással
többé-kevésbé szoros kapcsolatban állnak.
A fajok közti kölcsönhatások az egyik partner kipusztulását
eredményezhetik akár az erôs versengés, akár
a szimbionta partner elvesztése miatt. A zárva termô
növények evolúciós értelemben vett virágzásával
párhuzamosan csökkent az összes többi edényes
növény nemzetségeinek száma. John Maynard
Smith, angol biológus szerint sok szûznemzéssel
szaporodó élôlény sorsát az ivartalan
szaporodás és az abból fakadó kis változatosság
pecsételi meg. Egyesek szerint az ökoszisztémáknak
hosszú feltöltôdési és kisülési
idôszakai vannak. A kisülési idôszakok elôtt
az életközösség telítôdik fajokkal,
majd tömeges kihalás következik be. A bioszféra
belsô eredetû megváltozásai is vezethetnek fajok
kipusztulásához. A kezdetben reduktív légkört
az élôvilág tette oxidatívvá. Ez sok
olyan baktérium kihalását okozta, amelyek képtelenek
voltak eltûrni a magas oxigénszintet.
Az abiotikus tényezôk két csoportba oszthatók,
és elsôsorban a nagyobb kihalások hátterében
állhatnak. A földi okok közt említik a vulkáni
tevékenységet, a klímaváltozást, a tengerszintváltozást,
sôt még a kontinensvándorlást is. A tengerszint
süllyedése sok esetben egybeesett a tömeges kihalási
események idôpontjával. Sokszor viszont nincs semmilyen
összefüggés, emiatt a tengerszintváltozást
csak mint alkalmanként jelentôs hatást ismerik el.
Földön kívüli hatás lehet egy meteorit becsapódása,
vagy a Naprendszer áthaladása a Tejútrendszer egy-egy
karján. Utóbbi periódusideje megegyezik a kihalások
sokak által feltételezett periódusidejével.
A Földön kívüli okok Alvarez és munkatársainak
1980-as cikke nyomán robbantak be a köztudatba. A dinoszauruszok
hanyatlásának korából származó
kôzetekben szokatlanul nagy mennyiségû iridiumot mutattak
ki, és emellett szokatlan szerkezetû kvarcot. Mindkettô
ritka a Földön, de gyakori a világegyetem más részeibôl
származó kôzetekben. Ez az alapja annak az elméletnek,
mely szerint a dinoszauruszok és például sok hajtásos
növény kréta végi kihalását egy
becsapódó aszteroid okozta. A meteorit átmérôjét
10 kilométeresre becsülik, és feltételezik, hogy
a becsapódás után néhány évig
sötét, hideg porfelhôben kellett gondoskodni a megélhetésrôl.
A krízist viszonylag könnyen átvészelték
a páfrányok és az éjszakai életmódot
folytató melegvérû állatok, például
emlôs ôseink. A rendkívül izgalmas és népszerû
elméletet rengeteg adat alátámasztja. Ettôl
függetlenül a dinoszauruszok kihalásának magyarázatára
szinte divat elméleteket gyártani. Az ôshüllôk
tragikus sorsát magyarázzák a modern növényi
mérgek megjelenésével, a szemtelen emlôsök
tojásevô szokásával, a szén-dioxid szintjének
emelkedésével, a hímek fertilitási problémáival,
sôt krónikus pajzsmirigyproblémáikkal is.
Stephen Jay Gould a véletlen szerepét hangsúlyozza
általában az evolúció során is, és
a kihalási események alakulásában is. Evolúciós
folyamatokat szimuláló modelljei a tényleges kihalási
dinamikához hasonló, orsó alakú mintázatokat
mutatnak (4. ábra). Már Darwin korában is okozott
problémákat a véletlennek mint komoly evolúciós
tényezônek az elismerése, és ma is sokan idegenkednek
ettôl. Ennek ellenére általánosan elfogadott,
hogy legalábbis a tömeges kihalások alkalmával
sok múlik a szerencsén. Ha a kôzetlemezek mozgása
egy sziget eltûnését okozza, a csak ott élô
fajok akármilyen csodálatosan is alkalmazkodtak környezetükhöz,
többségükben elpusztulnak.
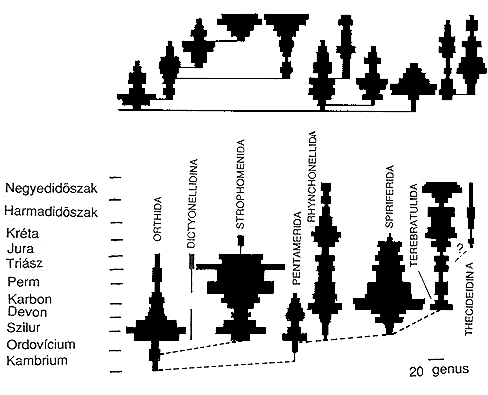
4. ábra. A pörgekarúak evolúciójában
a fajképzôdés és a kihalás dinamikája
jól köze(íthetô véletlenszerû modellek
elôrejelzéseivel. Az orsó alakú diagramok szélessége
a diverzitás mértékével azonos.
(Gould és munkatársai, 1997 nyomán, Vida, 1985)
Végül a legmodernebb típusú magyarázatokról.
A bioszféra bonyolult rendszer, és az ilyenek gyakran viselkednek
kaotikusan. A kaotikus rendszerek között vannak olyanok, amelyek
önszervezôdô jellegüknél fogva kritikus állapotok
felé tartanak. Ebben az esetben mindenféle méretû
katasztrófák történhetnek a rendszeren belül.
A katasztrófák mérete és gyakorisága
közt ismert az összefüggés, ez a hatványtörvény
(power law). A kihalások kutatásával foglalkozó
fizikusok modelljei szerint az ilyen rendszerek tulajdonságai sokban
hasonlítanak a biológiai rendszerekére. Az önszervezôdô
és kritikus állapotban lévô rendszerekben a
katasztrófák dinamikája jól megfeleltethetô
kihalási mintázatoknak. A megközelítés
túl elvont a biológusok többsége számára,
akik a modellek realitását kritizálják.
Az ember hatása
Az ember csak a pleisztocén óta fejt ki komoly hatást
a kihalási események alakulására, de "lemaradását
igyekszik gyorsan pótolni". Az egyes kontinenseken élô
nagy testû emlôsök és futómadarak kihalása
drámai módon függött az ember megjelenésétôl.
Ez alátámasztja azt a feltételezést, hogy az
emberiség bölcsôje Afrika volt. Az itteni nagy testû
állatvilág ugyanis sokkal kevésbé volt érzékeny
az ember jelenlétére, mint például az ausztráliai.
Ennek oka a hosszú közös evolúciós idôszak
volt, egymás mélyebb megismerése és megszokása.
Ma ezerszer-tízezerszer gyorsabban pusztulnak ki a fajok, mint
a megszokott háttérkihalás mértéke.
A mai tömeges kihalás egyik sajátossága az élôhelyek
erôteljes feldarabolódása. A gyorsan felaprózódó
élôhelyeken az élôlények sok, addig elônyös
tulajdonsága hátrányossá válik. Egy
vándorló állat minden tulajdonsága valamilyen
szinten összefügg a vándorlással. Ha a vándorlás
lehetôsége megszûnik, sok, addig elônyös
tulajdonsága a visszájára fordul és akár
a kipusztulását is okozhatja. Például ha egy
faj télen melegebb területekre költözött, nem
volt szüksége a téli bunda növesztésének
képességére. Ha az élôhely feldarabolódása
miatt nem vándorolhat, megfagy. Érthetô, hogy manapság
egyre nagyobb figyelem kíséri az ökológiai folyosókkal
kapcsolatos kutatásokat (lásd Gyulai Iván írását
a Természet Világa 1996-os, biodiverzitásával
foglalkozó különszámában). A változásokat
– a kutatók számára keserûen – jelzi, hogy az
ökológiai kutatásokban régebben alkalmazott terület/fajszám
összefüggés ma sokszor nem használható az
élôhelyek aprózódásának erôs
torzító hatása miatt. A kihalások legújabbkori
történetének forgatókönyvírója
az ember, de félô, hogy már fôszereplôjévé
is vált.
Összefoglalásul szeretném hangsúlyozni, hogy
a kihalás nem valamilyen tragikus, elkerülhetetlen esemény.
Természetes folyamat, a fajképzôdéshez hasonlóan
evolúciós mechanizmus. Emellett a bioszféra nem teljesen
elszigetelt rendszer, így Földön kívüli tényezôk
is beleszólhatnak az események alakulásába.
Ezek jelentôsége alkalmanként változó.
A kihalási folyamatokat elôidézô hatásoknak
egy része kiismerhetô. Ezek ellen a szervezetek védekezhetnek.
Ami az embert illeti, egyelôre még nem biztos, hogy a jelen
dinoszauruszai vagyunk. Csak tartunk afelé. A megoldást valószínûleg
nem a 21. század ragyogó biotechnológiája által
elôállitott fantasztikus élelmiszerek jelentik majd,
hanem a népesség létszámának nagymértékû
csökkenése. Ennek hiányában mi is bekerülhetünk
oda, ahol már úgyis nagy a tömeg: a Holocén Parkba.
IRODALOM
Alvarez et al. 1980 Extraterrestrial cause for the Cretaceous-Tertiary
extinction: experimental results and theoretical interpretation. Science.
208:1095–1108.
Benton, M. J. 1995 Diversification and extinction in the history of
life. Science. 268:52–58.
Hallam, A. & Wingall, P B. 1997 Mass Extinctions and their Aftermath.
Oxford University Press. Oxford.
Jablonski, D. 1986 Background and mass extinctions: the alterations
of macroevolutionary regimes. Science. 231:129–133.
Raup, D. M. 1986 Biological extinction in Earth history. Science. 231:1528–1533.
Raup, D. M. & Sepkoski, J. J. 1984 Periodicity of extinctions in
the geologic past. PNAS. 81:801–805.
Wilson, E. O. 1992 The Diversity of Life. Penguin Books. London.
| Természet Világa, |
129. évf. 3. sz. 1998. március, 105–108. o.
https://www.kfki.hu/chemonet/TermVil/
https://www.ch.bme.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez